 欢迎来到乐鱼网页版登录入口网站!欢迎来到乐鱼网页版登录入口网站!
欢迎来到乐鱼网页版登录入口网站!欢迎来到乐鱼网页版登录入口网站!
15221734409



本研究通过绘制小鼠24种组织的N-糖蛋白组全景图谱,揭示了糖基化既具有强烈的组织特异性(相同蛋白不同组织可携带不同聚糖),又遵循非随机的结构组合规律(同一糖基化位点上的聚糖共存受酶学偏好约束),两个核心发现共同指向糖基化是一种精确调控而非松散随机的修饰系统。
《Nature Communications》(简称 “Nat Commun”)是自然科研(Nature Research)旗下的国际顶级开放获取(Open Access)综合性学术期刊,2010年正式创刊,旨在发表自然科学领域(涵盖生物、化学、物理、地球科学、医学等)具有重要科学意义、但未达到《Nature》主刊突破性高度的原创研究成果,填补了顶级主刊与专业子刊间的发表空白。
出版周期: Bimonthly;
影响因子:2024-2025最新影响因子为15.7,五年影响因子为17.2;
ISSN:2041-1723;
发文量:2024 年发文量为10749篇;
版面费:$6790.00/篇;
一、研究背景
N-糖基化作为最重要的蛋白质翻译后修饰之一,在蛋白质折叠、细胞识别、信号传导和免疫应答等生物学过程中发挥关键调控作用,但由于聚糖结构的高度复杂性和微观不均一性(即同一糖基化位点可携带多种不同聚糖结构),系统性解析组织间糖蛋白组全貌一直面临重大技术挑战。尽管小鼠作为生物医学研究的核心模式生物,其基因组、转录组、蛋白质组和磷酸化修饰组已有较系统的组织图谱,但糖蛋白组的组织特异性景观长期缺乏高分辨率、大规模的系统性描绘——传统糖组学分析游离聚糖虽然能揭示部分组织差异,却丢失了聚糖所附着的蛋白载体和糖基化位点信息,导致功能解读受限;而现有的糖蛋白组学工具多依赖于预设聚糖组成数据库,难以区分具有不同生物学功能的同分异构体结构。因此,亟需建立一种能够同时解析聚糖精细结构、糖基化位点和蛋白载体信息的技术体系,从分子层面全面揭示N-糖基化在小鼠不同组织中的分布规律、功能意义及其调控机制。
二、关键技术总结
样本制备与糖肽富集技术
研究首先从24种小鼠组织(涵盖8大系统,含7个脑区及血清)中提取蛋白并酶解为肽段。为高效富集完整糖肽,采用亲水相互作用色谱(HILIC)与混合型阴离子交换色谱(MAX)两步串联策略,显著去除非糖肽干扰。对于22个组织,进一步通过高pH反相高效液相色谱将HILIC富集的糖肽分级为6个馏分,有效降低样本复杂度,大幅提升了糖肽鉴定的深度和覆盖度。
质谱数据采集与双碰撞能碎裂策略
富集后的糖肽经纳升液相色谱分离后,由Orbitrap Fusion Lumos高分辨质谱进行分析。核心技术创新在于对每个糖肽前体依次采用两种碰撞能进行数据依赖性采集:较高的HCD能量(33%)用于碎裂肽段骨架以鉴定氨基酸序列,而较低的HCD能量(20%)则用于保留并解析完整的聚糖结构信息。这种双碰撞能策略实现了在一针进样中同时获取肽段和聚糖两层信息,避免了分别进样带来的匹配误差,为后续结构解析提供了高质量谱图基础。
基于StrucGP的聚糖结构从头解析与鉴定
数据分析的核心工具为课题组自主开发的StrucGP软件。与传统工具依赖预设聚糖组成数据库不同,StrucGP利用低能谱图中特征性的B离子(聚糖碎片)和Y离子(肽段+部分聚糖碎片),采用模块化策略从头解析N-聚糖的精细结构,能够明确区分具有相同单糖组成但连接方式不同的同分异构体。结合UniProt小鼠蛋白数据库搜索,分别在肽段水平和聚糖水平均设定1%的假发现率(FDR),确保了鉴定结果的高可靠性。
机器学习驱动的置信度评估与生物信息学分析
为系统评价聚糖结构鉴定的可信度,研究构建了基于逻辑回归的置信度评估框架。以人工标注的774张谱图、472种聚糖作为金标准训练集,输入包括谱图匹配分数、GlyTouCan数据库匹配、聚糖类型概率、分支结构概率和生物合成路径合理性等5项特征,经交叉验证优化后为每个聚糖输出0–1的置信度分数。最终88.2%的聚糖被归为高置信度(≥0.82),仅高置信度数据用于后续生物学分析。在此基础上,研究进一步运用聚类分析、PCA降维、GO功能富集、糖基转移酶表达关联分析及共现网络构建等多种生物信息学手段,系统揭示了组织特异性糖基化模式及其调控规律。
研究通过“双碰撞能质谱碎裂 + StrucGP结构解析 + 机器学习置信度评估”三位一体的技术体系,实现了在不依赖预定义聚糖库的前提下高置信度地区分N-聚糖同分异构体,首次完成了小鼠多组织N-糖蛋白组的结构分辨率级系统绘制。
三、主要研究成果
1、24种小鼠组织N-糖蛋白组数据总览
通过对24种小鼠组织的195万余张含氧鎓离子特征谱图进行严格筛选与鉴定,在肽段水平1%假发现率(FDR)控制下,鉴定出17,806条含糖基化位点的肽段,对应13,934个N-糖基化位点和7,365个N-糖蛋白;进一步在聚糖水平同样以1% FDR过滤后,获得74,277个独特的完整糖肽,包含3,045种具有明确结构特征的N-聚糖,其中2,687种(88.2%)被归类为高置信度,后续分析主要基于此子集、8,681个糖基化位点和5,026个糖蛋白,构成了迄今规模最大的哺乳动物组织特异性糖蛋白组资源之一。在结构新颖性方面,鉴定的3,045种聚糖中有76.3%的组成和34.2%的精细结构已在国际聚糖结构数据库GlyTouCan中有过记录,说明本研究在聚糖结构层面贡献了大量新发现。为确保数据可靠性,作者建立了基于逻辑回归的整合置信度评估框架——以人工标注的472种聚糖(分属“结构合理”与“结构存疑”两类)为金标准训练集,模型表现出良好的判别能力(AUC=0.79,F1=0.70),并通过统计优化的阈值将88.2%(2,687种)的聚糖归为高置信度类别。在覆盖度方面,这些高置信度糖蛋白涵盖了UniProtKB中已审阅N-糖蛋白的63.5%,同时新发现了2,795个此前未在蛋白水平注释的糖蛋白;与MSFragger-Glyco、Byonic、pGlyco3等其他主流软件的比较显示各工具鉴定结果存在互补性,而基序分析确认了N-X-T序列偏好高于N-X-S的已知规律。这一系列数据表明,本研究所构建的图谱不仅在数量上实现突破,更在结构分辨率和鉴定可信度上达到了新的高度,为后续所有组织特异性分析和功能解读奠定了坚实的数据基础。
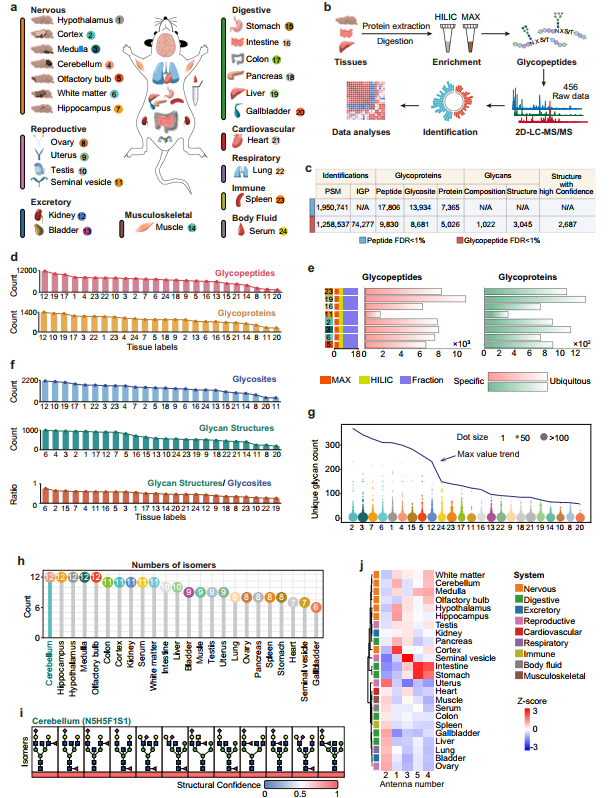
图1、24种小鼠组织的高分辨率N-糖蛋白组综合图谱
2、组织间糖基化鉴定数量的差异及其影响因素
研究人员从定量角度系统比较了24种小鼠组织在糖基化鉴定深度上的差异,并验证了数据集的可靠性与技术稳定性。在鉴定数量方面,各组织间呈现出巨大悬殊:胆囊因样本量限制未进行高pH分级,仅鉴定出1,275个完整糖肽和284个糖蛋白,而肾脏则高达11,723个糖肽和1,400个糖蛋白——对匹配组织的对比分析证实,分级处理可显著提高糖肽、糖蛋白、糖基化位点和聚糖各维度的鉴定数量,说明样本分级深度是影响鉴定覆盖度的重要因素。值得注意的是,即便在相同次数的液相色谱-串联质谱(LC-MS/MS)分析条件下,不同组织的鉴定数仍差异显著:肝脏的糖蛋白和糖肽鉴定数量最多,而精囊则最少,这一趋势与既往多组织蛋白质组学研究中蛋白丰度的组织间分布规律高度一致,提示组织固有的糖蛋白表达丰度差异是决定鉴定数量的内在因素。在数据可靠性验证方面,同一样品相同馏分的重复质谱分析显示,聚糖层面和糖基化位点层面的相关系数分别达到0.59–0.88和0.61–0.83,表明技术重复性良好;更为关键的是,组织间的变异幅度远大于技术重复间的波动,这有力地证明观察到的组织特异性糖基化模式并非由实验操作或仪器误差引起,而是反映了真实的生物学差异。综上所述,通过对鉴定深度的影响因素分析和技术重复性评估,为后续所有跨组织比较和生物学发现提供了坚实的数据质量保障。
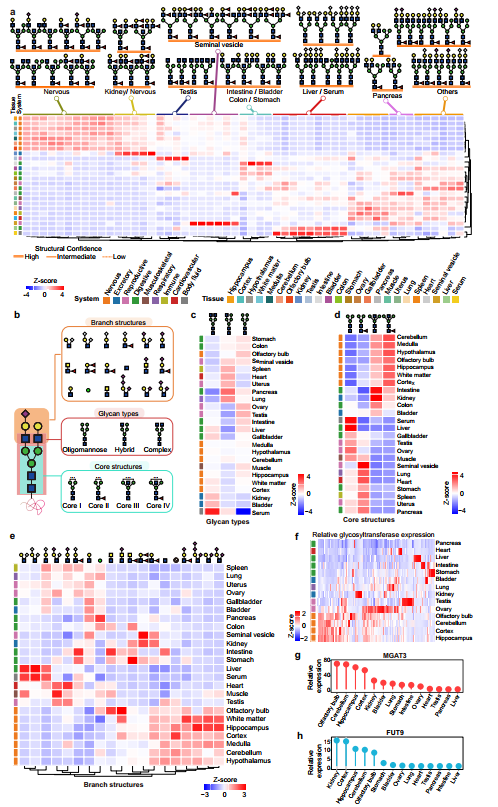
图2、24种小鼠组织中不同的聚糖结构模式与亚结构特征
3、组织间聚糖多样性与结构复杂度的差异模式
利用肾脏数据量大的优势,作者进一步探究了性别对糖基化的潜在影响;聚焦于不同小鼠组织在聚糖结构多样性与复杂度上的特征差异,揭示了组织特异性的糖基化规律。在糖基化位点数量上,肾脏以2,133个糖基化位点修饰以898种独特聚糖居首,这既受质谱分析深度影响,也反映了肾脏固有的糖蛋白丰富度;相比之下,神经系统七个脑区的糖基化位点数量中等(1,322至1,714个),却展现出最高的聚糖结构多样性(每脑区818至992种独特聚糖),其聚糖/糖基化位点比值高达0.54至0.75,远高于肾脏的0.42和肝脏的0.25,表明神经组织中每个糖基化位点平均携带更多种类的聚糖。尤其值得注意的是,神经组织中单个糖基化位点最多可携带257至367种不同聚糖结构,微观不均一性远超过其他组织。异构体分析进一步揭示了这一特征的组织偏好性——同一聚糖组成在不同组织中可扩展为6至12个结构异构体,其中神经系统含有最多的异构体,这些结构异构体可通过低能HCD谱图中特征性的B/Y离子以及C18色谱保留时间差异进行明确区分。在聚糖结构复杂度方面,神经系统聚糖以单天线和二天线为主(复杂度低),而二天线和三天线聚糖主导大多数组织,三天线和四天线聚糖则在胰腺和精囊等外分泌器官中尤为丰富,高分支(四天线及以上)聚糖多见于胃和肠组织。这些差异提示聚糖结构复杂度与组织的特定生理功能密切关联——神经系统偏好低复杂度但高多样性的聚糖可能服务于精细的神经信号调控,而外分泌器官和消化道的高分支聚糖则与其分泌功能或黏膜屏障保护需求相适应。综合而言,通过对聚糖数量、多样性和结构复杂度的多维比较,描绘了组织特异性糖基化的基本轮廓,为后续深入分析特定结构模块的分布规律奠定了基础。
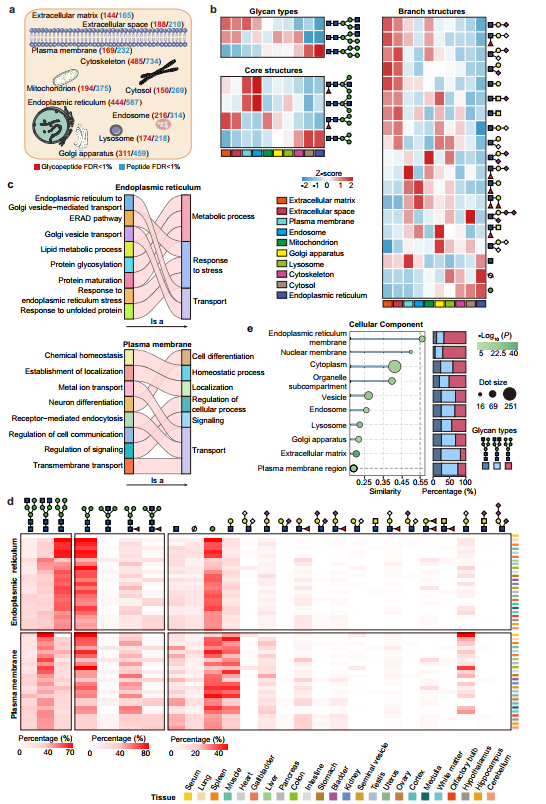
图3、24种小鼠组织中不同亚细胞定位间N-聚糖的保守性与多样性
4、不同组织中糖基化和糖蛋白的功能特异性
利用肾脏数据量大的优势,系统探究了雌雄小鼠之间的N-糖基化差异,揭示了性别对糖基化修饰的精细调控作用。在糖肽、糖基化位点和糖蛋白的总体水平上,雌雄两性高度相似,重叠率与相关性均表现良好,表明性别并非决定糖基化整体格局的主要因素。然而,在聚糖修饰的精细层面,差异依然清晰可辨:雌性肾脏明显富集含有LacdiNAc和Neu5Ac(N-乙酰神经氨酸)的聚糖结构,而雄性肾脏则呈现出更多Neu5Gc(N-羟乙酰神经氨酸,为小鼠特有的唾液酸类型)修饰的聚糖。功能富集分析进一步揭示了这些差异的潜在生物学意义——雌性富集的糖蛋白主要与免疫应答和生殖过程相关,而雄性富集的糖蛋白则倾向于参与代谢调控和细胞定位等基础功能。这些发现表明,尽管两性在糖基化整体特征上高度一致,性激素可能通过调控特定糖基转移酶的表达水平或催化活性,在聚糖的分支延伸和末端修饰层面产生细微但具有功能相关性的差异。值得强调的是,这些性别差异性聚糖修饰并非随机分布,而是与特定的生物学通路紧密关联,提示糖基化可能参与了性激素介导的器官功能调节。这为后续研究提供了重要启示:在进行组织糖基化研究时,性别是一个不可忽视的变量,尤其当研究聚焦于免疫、代谢和生殖相关功能时,雌雄差异可能对结论产生潜在影响。
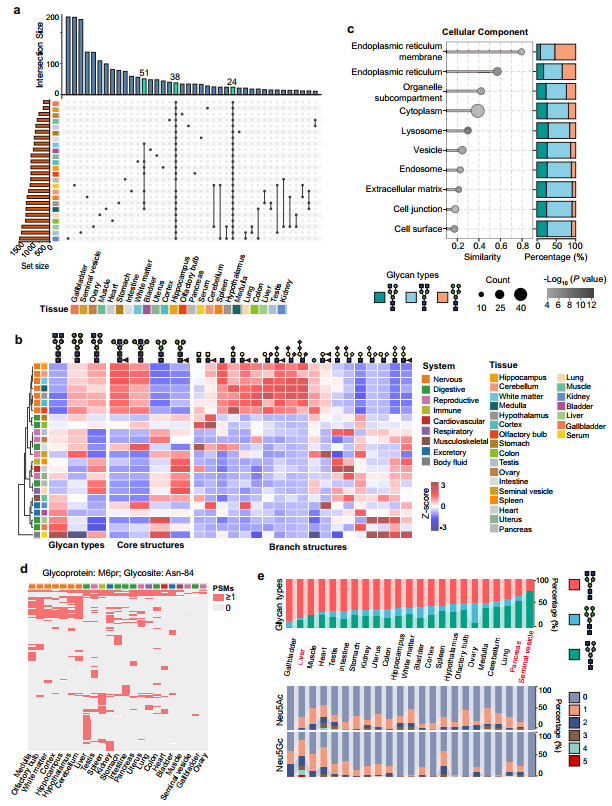
图4、不同组织中共同表达糖蛋白的聚糖结构比较
5、组织特异性聚糖结构特征及其酶学调控基础
结果展示了24种小鼠组织中最具代表性的高丰度聚糖结构,并揭示了其与糖基转移酶表达之间的内在关联。首先,作者提取各组织中按修饰糖基化位点数量排名前十的N-聚糖(排除常见的寡甘露糖型),发现不同组织呈现出鲜明的结构偏好:神经组织和肾脏共同富集核心岩藻糖基化的平分型聚糖;精囊虽与肾脏均有高水平Lewis结构,但平分型聚糖在精囊中极为罕见;肝脏和血清以低核心岩藻糖的唾液酸化复杂型为主;睾丸特异性富集末端GlcNAc;胰腺、肠、结肠和胃等消化器官共同富集末端半乳糖,但胰腺以杂合型为主,消化组织则携带平分型核心,精囊则以末端半乳糖的强岩藻糖基化为特征而不含平分型结构。为量化这些差异,作者将鉴定的聚糖分解为3种类型、4种核心结构和17种分支结构进行系统统计:在聚糖类型上,血清以复杂型占绝对优势(74.9%),睾丸以寡甘露糖型为主(38.1%),胰腺杂合型丰富(30.7%);在核心结构上,Core I在血清和肝脏中最高,Core II富集于精囊、心脏和肺,Core III在神经组织和消化器官中突出,而Core IV(岩藻糖化平分型)则特异性富集于脑区(平均29.4% vs 全组织13.1%);在分支结构上,神经系统以末端GlcNAc、Lewis表位和Neu5Ac为主(缺乏Neu5Gc),肝/血清以Neu5Gc为主,心脏/肌肉富集唾液酸化LacNAc,消化器官富集LacdiNAc,精囊高表达Lewis⁶表位。为探究这些结构差异的分子机制,作者利用公共RNA-seq数据绘制了209个糖基转移酶(GT)的组织表达热图,结果显示这些酶呈现明显的组织分辨表达模式。其中,Mgat3(合成平分型核心的酶)在脑区的高表达与Core III/IV的富集高度一致,Fut9(合成Lewis X的酶)在肾脏和脑区的高表达也与其相应聚糖的分布相吻合,从转录调控层面为组织特异性聚糖结构模式提供了直接的酶学解释。综上所述,通过对聚糖结构模块的组织分布统计和GT表达关联分析,建立起了从“聚糖结构差异”到“酶表达差异”的因果链条,为理解组织特异性糖基化的调控机制奠定了分子基础。
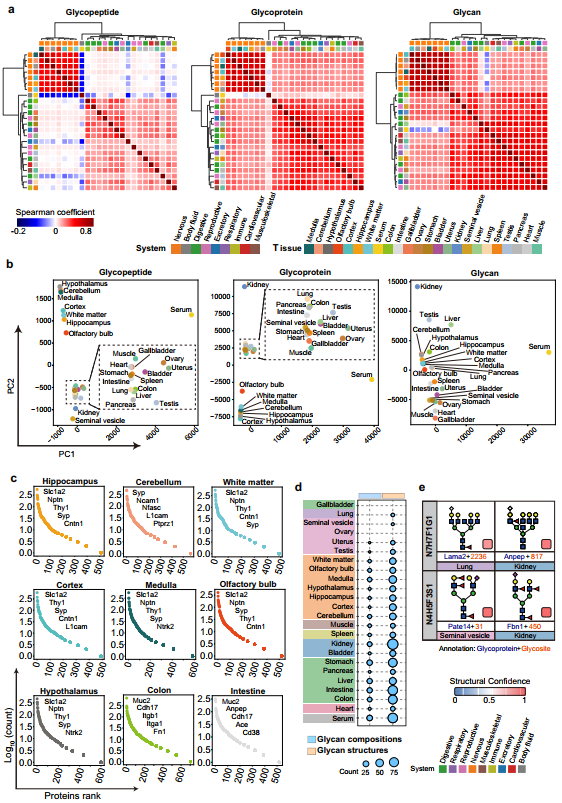
图5、位点特异性N-糖基化作为组织分化的分子标签
6、亚细胞定位对糖基化保守性与多样性的约束作用
随后,研究人员系统探讨了糖蛋白的亚细胞定位如何在其糖基化模式的保守性与多样性之间发挥关键约束作用。亚细胞定位分析显示鉴定到的糖蛋白主要分布于内质网(444个)、高尔基体(311个)和质膜(169个)。不同细胞器中的糖蛋白呈现截然不同的糖基化特征:内质网定位者主要携带寡甘露糖型简单聚糖,与N-糖基化在内质网启动并完成早期加工的合成路径一致;细胞外基质和质膜定位者则以复杂型聚糖为主,富含分支及末端唾液酸/岩藻糖修饰,代表高尔基体充分加工后的成熟状态。GO富集分析显示内质网糖蛋白主要参与蛋白折叠、质量控制和囊泡运输等保守功能,而质膜糖蛋白则执行信号转导、离子运输和细胞类型特异性过程等多样化功能,提示糖基化复杂度与功能多样性正相关。跨组织比较显示,尽管组织间差异存在,内质网和质膜各自的基本糖基化特征在所有组织中高度保守。选取在≥12个组织中共同表达的糖蛋白按亚细胞定位分组后计算组织间聚糖相似性,发现内质网定位者相似性最高(受保守合成途径限制),细胞外基质和质膜定位者相似性最低(适应特化胞外功能)。表明亚细胞定位通过在合成路径上的加工限制和功能需求上的选择压力,在糖基化的保守性与多样性调控中发挥决定性作用。
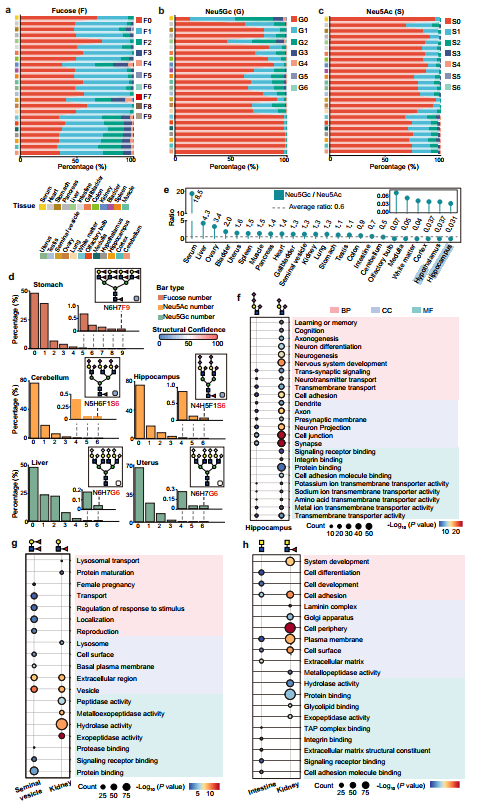
图6、24种小鼠组织中组织富集聚糖的功能分析
7、共同表达蛋白的组织特异性重塑及其组织身份判别能力
进一步的分析论证了位点特异性糖基化在组织功能适应性和组织身份识别中的核心地位。首先,即使同一蛋白骨架在不同组织中也经历系统的糖基化“改造”——在62个于至少12个组织中共同检测到的糖蛋白中,其附着的聚糖结构呈现显著的组织依赖性差异,且这种差异在24个通用表达蛋白中更为突出,仅分析这些共享蛋白的糖基化变体即可获得比全局聚糖分析更精准的组织聚类结果。以甘露糖-6-磷酸受体(M6pr)为例,其同一糖基化位点Asn-84在肝脏以高度唾液酸化的复杂型为主(延长蛋白半衰期、增强内吞回收),在胰腺以唾液酸化的杂合型为主,在精囊则以无唾液酸的杂合型为主,清晰展示了同一蛋白在不同生理环境中发生的“糖型开关”现象;但细胞表面透明质酸酶(Cemp2)的Asn-914位点在所有组织中保持高度保守,说明糖基化重塑是位点特异性的而非全或无现象。随后,在组织身份识别层面,通过无监督聚类和PCA分析证明,基于位点特异性完整糖肽的聚类能将24种组织按解剖系统和功能完美归类(脑区聚为一簇、血清独立分离、消化和生殖器官各自成簇),其区分度远优于基于糖蛋白整体或游离聚糖的聚类。作者还发现每个组织都拥有独特的“私有”聚糖结构,且同一聚糖组成的不同异构体在不同组织中富集模式截然不同——若仅分析组成则无法区分组织,只有引入异构体结构信息才能清晰鉴别。这些发现共同确立了“位点特异性糖基化”作为组织身份高精度分子标签的核心地位,揭示了糖基化通过在不改变氨基酸序列的前提下改变糖链结构来适配不同组织生理需求的调控逻辑。
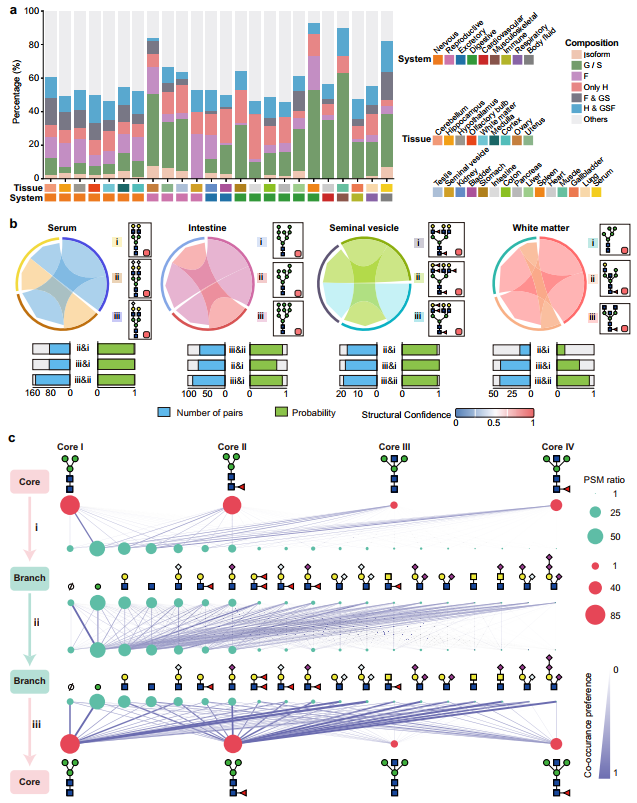
8、糖基化位点上的非随机聚糖共现规律
作者对每个具有至少10个聚糖观察值的糖基化位点进行了共现网络分析,发现绝大多数频繁共存的聚糖对之间的差异仅限于末端单糖:特别是仅相差1个唾液酸(S)或1个岩藻糖(F),而核心骨架(己糖H和N-乙酰葡萄糖胺G)保持不变。这种以唾液酸变异为主的共现模式在血清、肝脏、心脏、卵巢和肌肉等组织中尤为突出,二唾液酸化、单唾液酸化和去唾液酸化的二天线聚糖经常以高概率共存于同一糖基化位点,提示微观不均一性主要源于末端单糖的动态添加或去除,而非核心结构的剧烈改变。更为深入的亚结构共现分析揭示了核心与分支之间的偏好性配对规则:Core I主要搭配甘露糖分支,Core II偏向LacNAc和岩藻糖化分支,而Core III和Core IV(平分型核心)则倾向于与较少的分支结构共存。这一观察与已知的酶学抑制机制高度吻合——平分型GlcNAc可抑制GnTII、GnTIV、GnTV和FUT8等分支延长酶的活性,从而限制聚糖的进一步延伸。分支–分支共现模式同样显示出协调调控的特征:复杂分支常与简单分支(如单GlcNAc或LacNAc)共存,而polySia与Lewis结构的频繁共现则可能涉及细胞信号传导和免疫调节等协同功能。综合这些多层次网络分析,本研究得出核心结论:糖基化位点上的微观不均一性并非随机的谱图噪音,而是受糖基转移酶的底物偏好和结构约束共同塑造、具有高度组织规律性的有序组合系统。
四、研究的意义及不足
本研究首次绘制了小鼠24种组织的高分辨率N-糖蛋白组图谱,在数据规模、结构解析深度和鉴定可信度上均实现突破,填补了系统性糖蛋白组研究的空白。核心发现包括:糖基化具有强烈组织特异性,富集聚糖结构与各组织生理功能密切相关;亚细胞定位对糖基化多样性与保守性起关键约束作用;同一蛋白骨架可发生组织依赖性“糖型开关”,且糖肽在区分组织身份方面远优于蛋白质或游离聚糖。该资源为理解糖基化在组织分化、蛋白功能微调中的作用提供了系统性框架,对生物标志物发现和药物开发具有推动作用。然而研究也有一些不足之处,包括:样本混合策略掩盖个体差异,质谱深度在部分组织不一致,未覆盖硫酸化等罕见修饰,缺乏功能验证实验,且小鼠与人存在Neu5Gc等物种差异,限制了直接临床转化。未来需通过单细胞/空间糖蛋白组学、跨物种比较和疾病模型扩展进一步深化研究。
参考文献:
Wu, Y., Yang, M., Xu, Y. et al. A comprehensive N-glycoproteome atlas reveals tissue-specific glycan remodeling but non-random structural microheterogeneities. Nat Commun 17, 1448 (2026). https://doi.org/10.1038/s41467-025-68186-2

