 欢迎来到乐鱼网页版登录入口网站!欢迎来到乐鱼网页版登录入口网站!
欢迎来到乐鱼网页版登录入口网站!欢迎来到乐鱼网页版登录入口网站!
15221734409



英文名:Chemoproteomic profiling reveals histone H4 dopaminylation inhibiting cell growth
中文名:组蛋白 H4 多巴胺化(H4Q27dop)抑制神经母细胞瘤细胞生长
期刊:Nature Chemical Biology
年份:2026
作者开发了炔基化多巴胺探针,系统捕获活细胞中的多巴胺化蛋白,并发现 H4Q27dop 是一种具有转录抑制功能的组蛋白修饰:它削弱 CEBPD 对 CCND1 启动子的结合,降低 cyclin D1,导致神经母细胞瘤细胞 G0/G1 阻滞和增殖抑制。
研究类型:
化学生物学 + 表观遗传 + 肿瘤细胞生物学
核心贡献:
1.建立“活细胞多巴胺化蛋白”化学蛋白组学富集方法。
2.提出 H4Q27dop 是不同于 H3Q5dop 的转录抑制型标记。
3.给出DA–TGM2–H4Q27dop–CEBPD–CCND1–细胞周期 机制链。
为什么多巴胺化值得研究?
单胺类神经递质不只是配体信号,也可能直接成为蛋白修饰基团。
已知事实:多巴胺可通过 TGM 催化,共价连接到蛋白谷氨酰胺(Gln)侧链。H3Q5dop 已与成瘾相关转录可塑性有关。
关键缺口:缺少全局底物图谱;泛多巴胺化抗体工具有限;MDC 等通用单胺探针特异性不足。
本文问题:
能否用小分子探针在活细胞中捕获 DA 修饰底物?新位点是否具有疾病功能?
本文把神经递质—翻译后修饰—染色质调控—细胞周期串成了一条机制链。

总体研究逻辑
先找底物,再证功能,最后解机制。
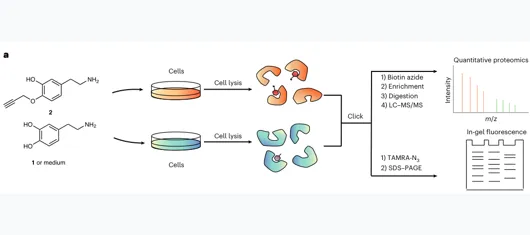
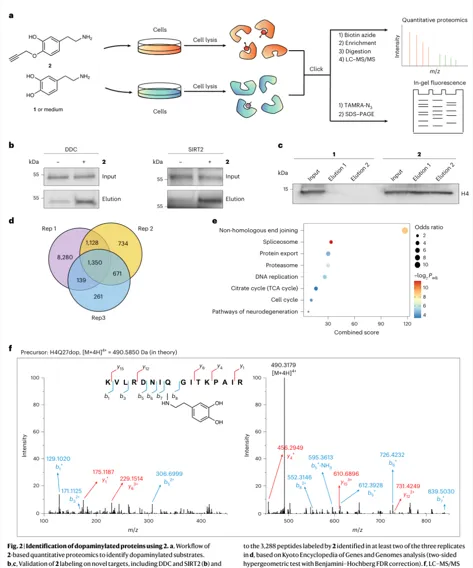
1.探针开发:propargyl dopamine(2)+ CuAAC click reaction
2.全局鉴定:Streptavidin富集+ LC–MS/MS / 肽段层面
3.位点验证:H4Q27dop:MS、抗体、TGM2依赖
4.功能验证:H4WT vs H4Q27A增殖、细胞周期、CCND1
5.机制闭环:ChIP-seq/RNA-seq/ATAC-MS CEBPD结合受阻
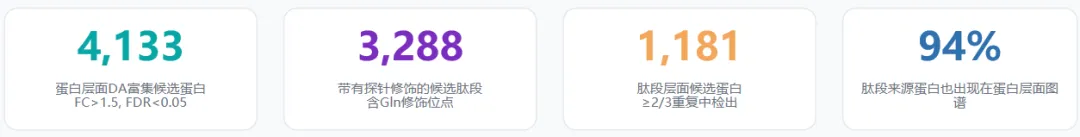
结果链条:
全局资源:4,133个DA富集蛋白;1,181个肽段层面候选蛋白
关键位点:H4Q27dop 被内源性MS与特异性抗体确认
机制模型:H4Q27dop↑ → CEBPD占位↓ → CCND1↓ → G0/G1阻滞
技术创新
炔基化多巴胺探针让DA修饰可点击、可富集、可成像
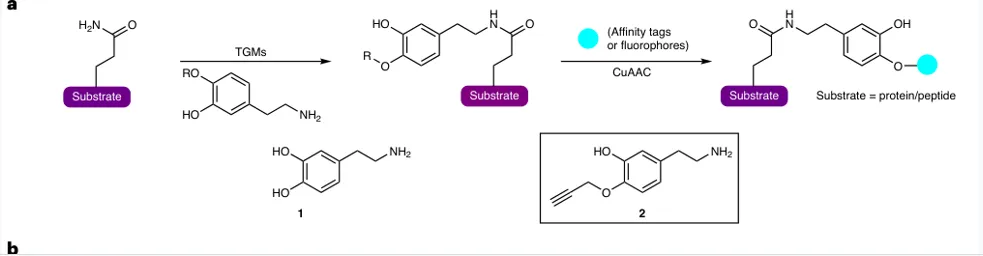
设计原则:在不参与多巴胺化反应的 4-hydroxyl 位置引入 terminal alkyne;保留被TGM2识别和连接的反应特征。
验证逻辑:体外TGM2酶促反应证明探针可连接到H3Q5;细胞内TAMRA成像显示核/胞质均有信号。
关键条件:SK-N-SH细胞:200 μM 探针处理2 h,用于后续蛋白组富集;≤500 μM未见显著毒性。
探针性能
剂量/时间依赖标记,并进入细胞核
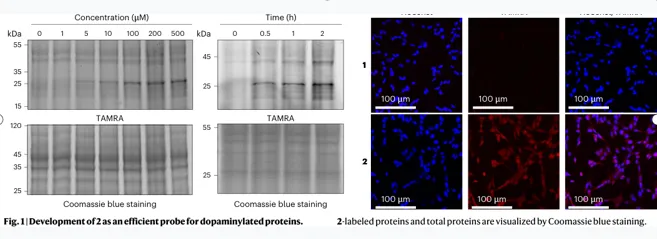
如何证明它有效?
TAMRA-azide点击后可见全蛋白荧光条带
200 μM / 2 h 是后续组学分析的优化条件
多巴胺竞争、TGM2敲低或抑制均降低信号共聚焦显示标记蛋白分布于核与胞质
评价:这一步是整篇文章的技术地基。它不是直接用抗体捞未知修饰,而是把多巴胺改造成可点击的化学报告分子,再通过生物素富集进入质谱。
全局图谱
从蛋白层面到肽段层面定位候选多巴胺化底物
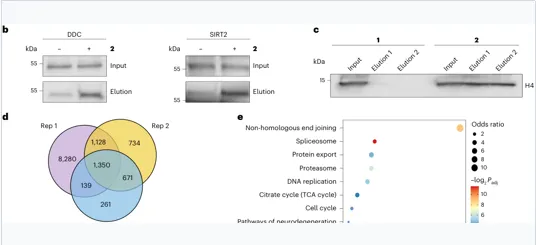
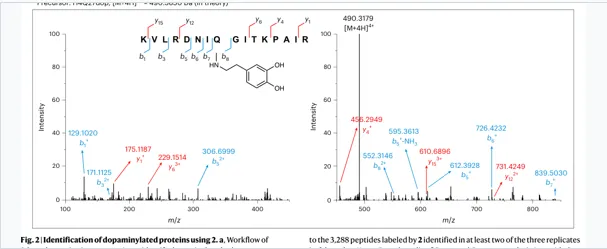
验证靶点:DDC、SIRT2、Histone H4;内源性组蛋白MS确认 H4Q27dop。
富集蛋白提示
多巴胺化可能广泛参与细胞周期与DNA相关过程
富集通路:
DNA replication|Cell cycle|Proteasome|Protein export|Spliceosome
为什么聚焦H4?
组蛋白H4位于细胞核,H4Q27处于N端尾部;结合组学富集、肽段MS证据与肿瘤细胞周期表型,具备机制深入价值。
从资源到机制:
4,133个候选蛋白构成资源库;但文章真正的高分点是从资源库中抽出H4Q27dop并完成“位点—转录—表型”的闭环。
功能验证
H4Q27dop 是多巴胺抑制细胞增殖的重要介质
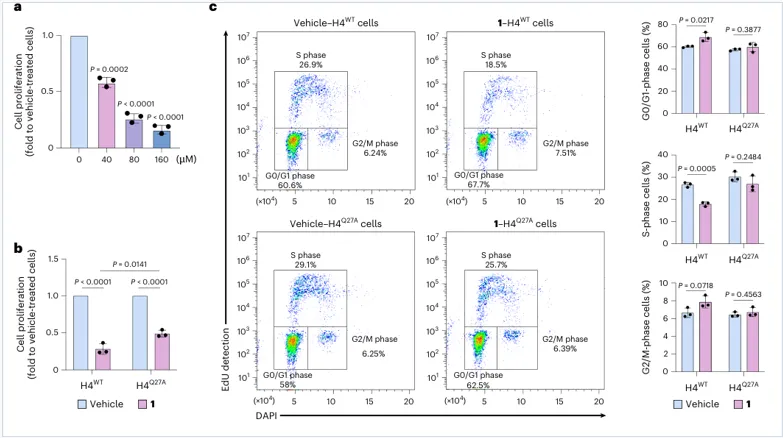
关键对照:
H4WT vs H4Q27A
Q27A使多巴胺诱导的增殖抑制和G0/G1阻滞明显减弱。
解释:
多巴胺对细胞周期的影响不是单纯毒性,而依赖H4第27位谷氨酰胺可被多巴胺化。
表型:
BrdU下降 + S期比例下降 + G0/G1比例上升
位点证据
多巴胺处理提高H4Q27dop,Q27突变削弱该修饰
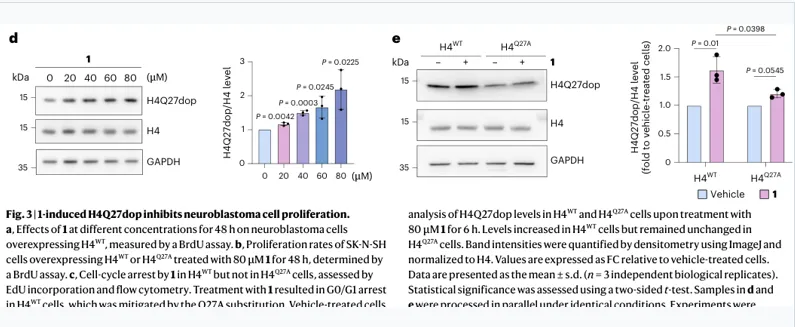
证据链:
构建H4Q27dop特异性兔单抗
TGM2敲低/抑制降低H4Q27dop
多巴胺剂量依赖升高H4Q27dop
H4Q27A细胞中该升高显著减弱
不只证明“多巴胺抑制增殖”,而是证明“多巴胺 → H4Q27dop增加 → 功能表型”的因果关系。
ChIP-seq显示
H4Q27dop广泛分布,且与较低转录活性相关
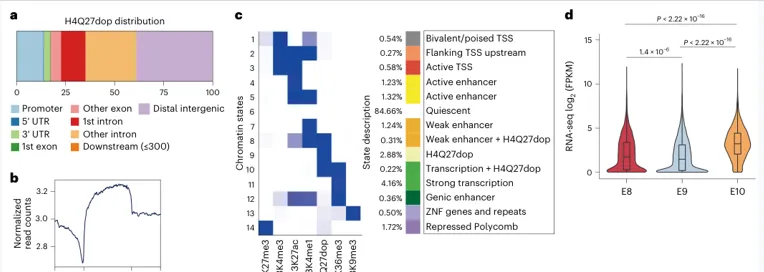
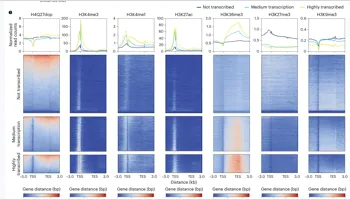
关键读数:177,593个H4Q27dop peaks;覆盖约2.44%基因组;约61.13%位于基因区域;H4Q27dop-only状态对应较低转录水平。
靶基因闭环
H4Q27dop升高伴随CCND1转录和Cyclin D1下降
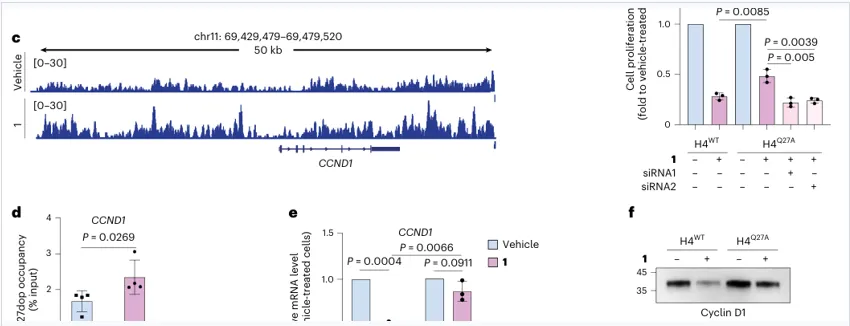
为什么是CCND1?
CCND1编码cyclin D1,是G1/S转换关键因子,正好解释多巴胺处理后G0/G1阻滞和S期减少。
H4WT中下降明显
H4Q27A中下降被削弱
在H4Q27A细胞中敲低CCND1,可恢复多巴胺诱导的增殖抑制,说明CCND1是该通路的关键功能输出。
机制关键
H4Q27dop阻碍CEBPD结合CCND1启动子
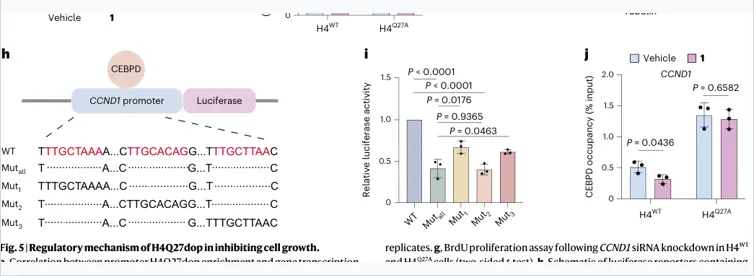
证据组合:
ATAC–MS筛选到CEBPD/NFXL1
CEBPD在SK-N-SH中表达更高
Luciferase证明CEBPD可激活CCND1启动子
ChIP–qPCR:多巴胺降低CEBPD占位,仅发生于H4WT
模型解释:
多巴胺提高CCND1区域H4Q27dop后,并不是简单“关闭染色质”,而是减少CEBPD对CCND1启动子的有效结合,从而降低CCND1转录。
整合模型
DA–TGM2–H4Q27dop–CEBPD–CCND1轴

化学蛋白组学证据:探针标记、富集、MS鉴定H4Q27dop位点确认
表观基因组证据:H4Q27dop ChIP-seq
与低转录活性相关
功能闭环证据:H4Q27突变、CCND1敲低CEBPD占位验证
H4Q27dop不是“被动标记”,而是可改变转录因子占位并产生细胞周期效应的功能性组蛋白修饰。
创新点
1. 工具创新
从“没有合适抗体/探针”切入,建立活细胞多巴胺化蛋白的化学蛋白组学平台。
2. 资源价值
给出大规模候选底物和肽段资源,为多巴胺化、单胺化研究提供可复用数据集。
3. 新修饰位点
H4Q27dop是新的功能性组蛋白多巴胺化位点,且与H3Q5dop方向不同。
4. 机制闭环
从探针→MS→抗体→ChIP-seq/RNA-seq→突变→TF结合→CCND1救援,证据链完整。
5. 疾病场景
选择神经母细胞瘤,与多巴胺代谢和儿童肿瘤表观遗传异常高度相关。
6. 概念扩展
提示神经递质可作为代谢物来源的PTM信号,直接调控染色质和细胞命运。
局限性与可借鉴方向
主要局限:
探针富集代表“可被探针标记的DA相关底物”,并非所有内源性多巴胺化位点都已明确。
大量候选蛋白仍未逐一验证;SIRT2内源性修饰位点也因丰度问题未定位。
功能机制主要在SK-N-SH模型中完成,缺少动物肿瘤/临床样本层面的验证。
多巴胺化的eraser、reader以及与其他组蛋白修饰的互作仍不清楚。
对课题设计的启发:
做新型PTM文章,要有“组学发现 + 位点抗体/质谱验证 + 突变体因果验证”。
机制层面最好从“修饰改变蛋白互作/TF结合/染色质状态”切入,而不是只检测表达量。
疾病场景要选择与代谢物来源高度匹配的模型,例如神经递质、乳酸、丙酮酸、脂肪酸等。
若要转化为医学文章,可补充组织样本、动物模型和治疗干预,增强疾病相关性。
Take-home message:
新型蛋白修饰研究的高分路径 = 代谢物/信号分子来源 + 化学/组学工具 + 精确位点验证 + 因果突变救援 + 疾病机制闭环。

